This is my fourth post on the World War I battlefields where my grandfather Will Mathers (1893-1953) served from 1916 to1918. See here, here, here, and here for previous posts.
After Flanders, Tom and Will spent December 2017 until March 2018 well behind the front lines near Boulogne-sur-mer in the Pas-de-Calais. They were sent to the front at Villers-Bretonneux in late March 1918.
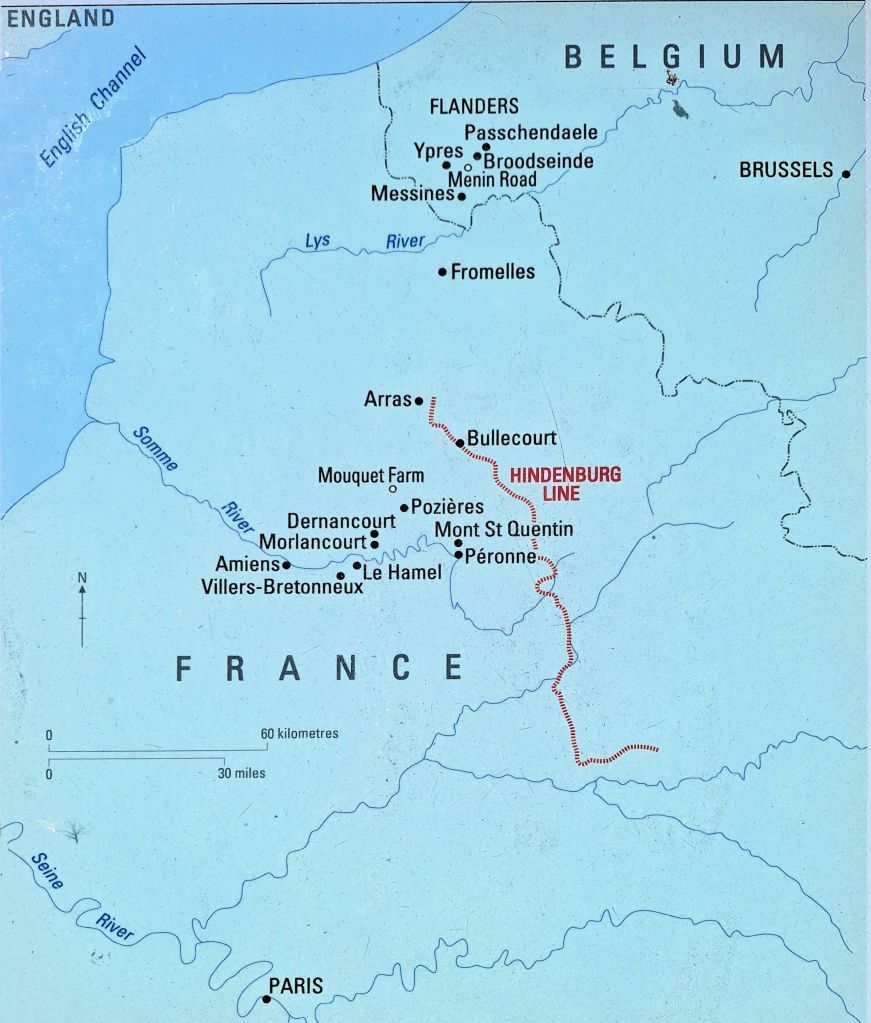
The Western Front – Flanders and the Somme. The Germans pushed the front line to Le Hamel and Villers-Bretonneux in March-April 1918
On 21 March 1918, the Germans launched an offensive against Allied forces around the Somme. Within two weeks they had advanced around 65 km, but were halted at Villers-Bretonneux by British and Australian troops on 4 and 5 April 1918. On 24 April, a renewed German attack captured the town, but that night the 13th and 15th Australian Brigades counter attacked from the north and south, driving the Germans from the town and nearby woodland. These battles were fought across the ground on which the Villers-Bretonneux Military Cemetery now stands.
Villers-Bretonneux Military Cemetery Villers-Bretonneux Military Cemetery with the Australian National Memorial in the background
The Cemetery contains more than 2100 graves of Commonwealth servicemen of the First World War, of whom more than 600 remain unidentified. At the back of the cemetery is the Australian National Memorial, which honours the Australian soldiers who served in France and Belgium. It consists of a central tower, with spectacular views of the surrounding countryside, flanked by wing walls commemorating the 10,796 Australian who died in France and who have no known grave. Along the upper edges of the Memorial, the battle honours of major engagements involving Australian troops in France and Belgium are inscribed.
The Australian National Memorial at Villers-Bretonneux The central tower of the Australian National MemorialThe view west over the Cemetery from the top of the Australian National Memorial tower.The view south towards Villers-Bretonneux from the Memorial TowerLooking down the Tower staircase
On 21 April, 1918 the 8th Australian Field Ambulance, including Will and Tom, were sent to relieve the 15th Australian Field Ambulance at Hamelet, a village a few kilometres north-east of Villers-Bretonneux. The German air ace Baron von Richthofen (the Red Baron) was shot down nearby on that day. The field where he crashed is across the road from the Australia Corps Memorial (see further below).
The Red Baron was the top German air ice of the First World War. With some 80 victories to his name, he shot down more aircraft in the war than any other pilot. His name re refers to his red Fokker triplane. His squadron was known as the Flying Circus, many decades later inspiring the name Monty Python’s Flying Circus. On the morning of 21st of April 1918, the red Baron was pursuing a British aircraft, and had another Canadian aircraft on his tail. He flew extremely low and was killed by machine gun fire from the 24th Australian Machine Gun Company.
The field where the Red Baron was shot down, seen from the Australia Corps Memorial
Two days later, Will and Tom were both gassed on the battlefield, and Will and Tom were gassed a second time on 30 April, suffering gas burns all over their bodies. After a couple of weeks to recover, they manned an aid post at Villers-Betonneux, carrying men back to the post from the front line under heavy shelling.
The Australia Corps Memorial is on a slight rise to the east of the village of Le Hamel. The memorial honours all those who served with the AIF in France and Belgium and bears the names of nearly 11,000 of their number who fell in France and have known no one grave. Around 3800 of those named here were killed during the battle of the Somme in July and August 1916. Another further 2800 fell in April and May 1917 at Bullecourt (Battle of the Hindenburg Line) and more than 2100 were lost in 1918 in the Battle of Le Hamel.
Australia Corps Memorial near Le HamelQuotation of French Prime Minister Georges Clemenceau on side panel of the Memorial
The Australia Corps Memorial is located at a section of German trenches, known to them as Wolfsburg, captured by the Australians early in the morning on 4 July 1918. The Australians occupied these trenches for about five weeks until they attacked from here in the great Battle of Le Hamel on 8 August 1918.
German trenches still visible near the Australia Corps Memorial
At 4:20 am on 8 August 1918, around 50,000 Commonwealth, French and American troops began a surprise assault on German lines to the east of armies. To give the newly arrived American Expeditionary Force (AEF) combat experience, the five Australian infantry brigades involved were augmented by four companies from US Army battalions. Hamel was the first time during the war that American soldiers were commanded operationally by non-American officers.
Attacking through dense fog they were well equipped with Lewis guns, trench mortars and rifle launched grenades. A creeping artillery barrage helped to shield the infantry, while enemy guns were silenced by gas and explosive shells. Assisting the assault troops were more than 550 armoured vehicles, including new Mark 5 and fast Whippet tanks which had moved into position in secret, their engine noise disguised by aircraft flying overhead. The Germans assumed the planes were trying to give them a sleepless night but had no clue about the coming attack.
Wounded Australians of the 15th Brigade, and wounded German prisoners sheltered beside a British Tank on August 9th, 1918. Australian War Memorial E02880.
General Sir John Monash (1865-1931) was an Australian civil engineer who took part in the Gallipolo campaign and in May 1918 became commander of the Australia Corps, at that time the largest corps on the Western Front. Monash planned a combined campaign of infantry and armoured vehicles, including 60 combat and four supply tanks. The plan called for a creeping barrage in which the artillery barrage moves slowly in front of the advancing troops, protecting them by keeping the defenders under cover. Monash was adamant that infantry should not be sacrificed in an unprotected advance. The objectives were all achieved in 93 minutes, three minutes longer than Monash’s calculated battle time. The Battle of Le Hamel has since become famous as a model for a completely successful all-arms attack.
The Allies consolidated their new positions during the day and advanced another 300 metres during the night. In the morning of 5 August, they took another 700 German prisoners. The Germans counter-attacked at 22:00 that night supported by phosgene and mustard gas bombardments. At 2 am on 6 July, the two Australian battalions and the attached Americans counter-attacked and overwhelmed the experienced German storm-troopers with grenades and clubs. The shocked Germans were forced back, restoring the Allied line and releasing captured Australian stretcher-bearers.
The Wolfburg trenches close to the Australia Corps Memorial
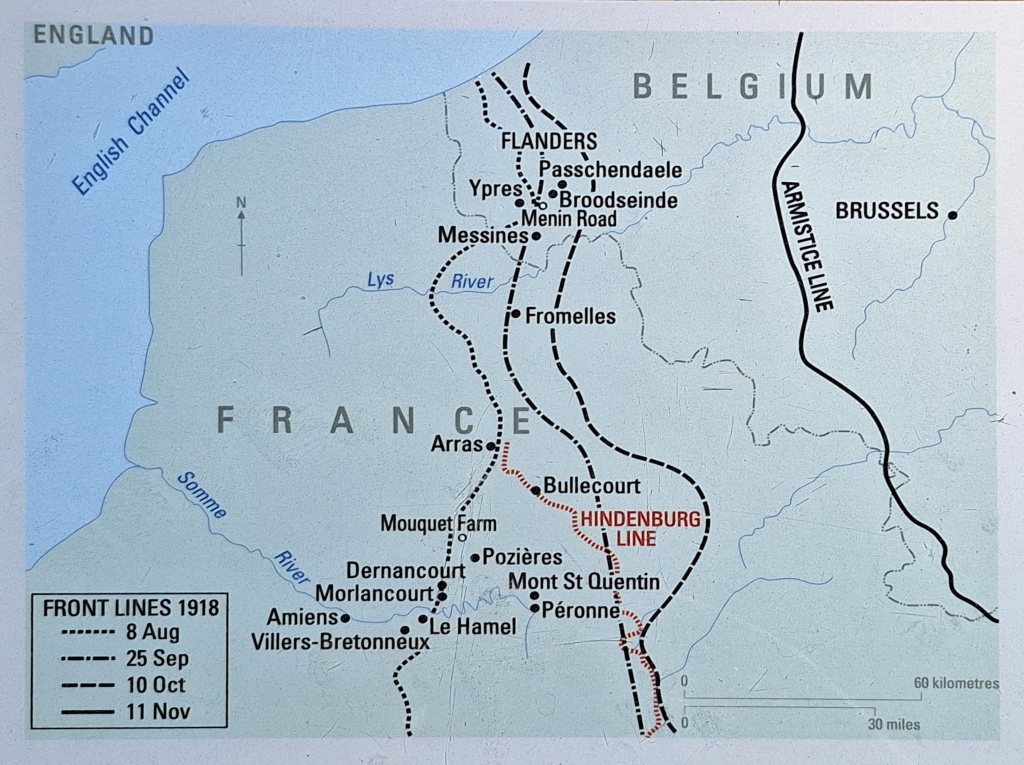
Over 4 days of fighting the Allies advanced as far as 13 kilometres in places. Some 20,000 Commonwealth soldiers were killed or wounded. The French army suffered around 24,000 casualties. Total German casualties may have been as high as 75,000, including more than 30,000 taken prisoner. Over the following weeks, a series of hard fought battles took place over the plains and uplands of the Somme, and by mid September, the Allies had recovered all the territory they had lost in March. The advance begun here did not cease until the Armistice.
From 5 August to 12 September 1918, Will and Tom followed the fighting on the front line, from Mericourt and Raincourt to Le Hamel and on to Péronne. They slept where they could, in a field, in a trench, in sump holes. There were thousands of prisoners and booty. On 4 September, Will was at Péronne and was wounded in the leg by shrapnel but remained on duty. On 10 September they were relieved by 13th Australian Field Ambulance and marched out.
For the two days we spent on the Somme, we stayed in the centre of the Old Town of Péronne, a 100 metres from the Chateau de Péronne, now housing a major museum of the First World War. On our second day, we spent a couple of hours in the museum, well worth a visit. The next morning we headed north back to German-occupied Belgium.
The main entrance to the Chateau de Péronne, flying the flags of the Allied nations and the EU.
From Flanders we headed south across the French border to the WW1 battlefields of the Somme. For my previous posts on Flanders, see here and here. The Somme offensive commenced at 7.28 am 1 July 1916 with the explosion of a massive mine detonated beneath the German front line. The Lochnagar Crater is the largest of the still extant mine craters of the Western Front.
Lochnagar Crater
The Lochnagar mine was the largest of 19 mines, placed beneath the German front line to assist the British infantry advance and left a crater 21 m deep and 100 m wide, which was captured and held by British troops. While the blast itself killed relatively few Germans, the vast majority of deaths occurred during the following infantry assault.
Despite their colossal size, the Lochnagar and other mines failed sufficiently to neutralise the German defences in La Boisselle. The ruined village was meant to be captured in 20 minutes but by the end of the first day on the Somme, the British 3rd Corps divisions had suffered more than 11,000 casualties for no result.
The Vimy Memorial
The spectacular Vimy Memorial sits at the highest point of Vimy Ridge, about eight kilometres northeast of Arras. It is a memorial to all Canadians who served their country in battle during the First World War, and particularly to the 60,000 who gave their lives in France. It also bears the names of 11,000 Canadian servicemen who died in France – many of them in the fight for Vimy Ridge – who have no known grave.
The figure of a cloaked young woman stands on top and at the centre of the front wall of the memorial and overlooks the Douai Plains. She has her head bowed, her eyes cast down, and her chin resting in one hand. The saddened figure of Canada Bereft, also known as Mother Canada, is a national personification of the young nation of Canada, mourning her dead.
Canada Bereft
The two pyramid-shaped hills in the distance are giant cone-shaped slag heaps, made of leftover rock from coal mining. When Felix saw them, he insisted we must climb them on our return trip to Belgium.
The ridge is pockmarked with mine and artillery craters. Many undetonated shells remain in the ground, and affected areas are fenced off with signs saying “No Entry – Undetonated explosives”
The ridge was a heavily fortified 7-kilometer hill held by the Germans. After previous French and British armies suffered around 150,000 casualties trying to take it, the Canadian Corps extensively planned their assault. They used aerial reconnaissance to construct a replica of the German trenches and bunkers, behind their front line. They trained extensively for weeks to develop an effective plan of battle. Unlike the British practice of briefing only officers on battle plans, the Canadians briefed every soldier, and they all knew what they had to do, irrespective of whether their officers were still leading them.
The Battle of Vimy Ridge (April 9–12, 1917) was one of the major battles in the Somme, where four divisions of the Canadian Corps fought together for the first time. This tactical victory against fortified German positions is considered a defining moment that forged a distinct Canadian national identity.
Part of the Canadian front-line trenchesIn the German front-line trenches only about 20 m from the Canadian trenchesA machine-gun emplacement in the German trenches
The Canadians extended a network of front-line trenches and tunnels constructed by the British. The tunnels were used to move troops, to connect underground dugouts and living quarters and for messengers to safely travel to and from the front lines. At Vimy Ridge, there were six kilometres of tunnels, and then longest tunnel was 1.6 km.
In one of the longer Canadian tunnels
Two days later, as we headed back from the Somme to Belgium, we did a slight detour to visit the giant slag heaps of Loos-en-Gohelle, located about 12 km northwest of Vimy. These two slag heaps are the tallest artificial mountains in Europe. We climbed the highest one, to the summit 146 m above ground level. It is 2 metres taller than the second slag heap.
Ascending Europe’s tallest slag heap, second slag heap partly visible on left.Approaching the summit.On the summit. Slag heap tourism hits new heights in FranceView south from the summit towards Vimy Ridge. The Canadian Memorial is visible in the middle close to the horizon,
This is my second post on the World War I battlefields where my grandfather Will Mathers (1893-1953) served from 1916-1919 (the first is here). After recovering from being gassed, and then some leave, Will and Tom went to the Messines Front on 20 November 1917 and ran a dressing station at Bailleul until mid December. Bailleul was 13 km west of Mesen, in French Flanders, just 3 km from the Belgian border.
The Messines Ridge extends south from Ypres almost to the Belgian border and was captured by German forces in 1914. The ridge granted the German Army a dominant position overlooking the Ypres salient. Before launching their offensive at Passchendaele in September 1917, the Allies planned to capture the Messines Ridge.
The battle of Messines, at Wijtschate and Mesen, was notable for two things. The Allied attack was preceded by the detonation of a million pounds of explosives packed into 21 mineshafts under the German positions along the entire 15km length of the Messines ridge. The colossal explosion in the early hours of Thursday 7 June 1917 was heard as far distant as London (where Lloyd George is reputed to have heard it from his offices in Downing Street) and Dublin.
Troops on the rim of one of the Messines mine craters, probably Peckham, shortly after the battle.
The two most northerly positioned mines on the Messines ridge exploded on Hill 60 (24,000 kg of explosives) and at the Caterpillar (32,000 kg of explosives), which is just across the railway tracks from Hill 60. The tunnel dug to place the explosives was 700 metres long. The mines left deep and wide craters that are there still today.
Hill 60 CraterCaterplillar Crater
. Although 10,000 casualties were reported on 10 June, 21 days after the explosion, the historian, Simon Jones, challenged the death toll of the mines using primary sources and suggested that the mine explosions killed no more than 500 German troops and that 7,344 were taken prisoner. The report of 10,000 casualties originated from the German estimates for total casualties (including prisoners) in the Battle of Messines.
Messines Ridge British Cemetery at Mesen in Flanders
The battle, and the taking of the town of Mesen (Messines in French) on a very hot summer day, was one of the few British and Allied ‘triumphs’ in Flanders. Although it was successful, a meagre advance was tempered by the loss, among the Allied troops, of 6800 Australians. By the end of the day Private John Melrose (aged 29) was among the dead. He is remembered on the Ypres memorial (panel 127) at the Menin Gate. Shortly afterwards the 36th was in action again, this time at Passchendaele.
Menin Gate, YpresPanel 127 on the Menin Gate, Melrose J recorded lower right column,
John Melrose (1887-1917) and William Melrose (1989-1917) were cousins of Will and Tom, who had emigrated to Australia in 1909 and 1911 respectively. They both enlisted in 1916, John only two weeks after he married Annie Lennox, another young immigrant from Scotland.
Will and John Melrose
John joined the 36th Australian Infantry Battalion, nicknamed “Carmichael’s Thousand,”and took part in the Battle of Messines. Assigned to the 9th Brigade, the battalion fought on the Oostaverne Line and suffered heavily.
The Red Cross wrote several letters to Annie. In a letter dated 1 March 1918, the report of a Private Jeffery states:
“I was personally acquainted with No. 2611 Pte Melrose,J. 36th Battalion. On the night of 7th June 1917 Pte. Melrose and I were part of a carrying party working between Ploegstreet Wood and firing line. About half way towards the firing line on our fourth trip an enemy machine gun opened fire. No. 2611 Pte. Melrose was hit in the head and killed. He was buried next morning about 100 yards from where he fell.”
John must have survived the attack on Messines Ridge (they started in the early hours) only to be killed when the victory was secured. Ploegsteert Wood was at the extreme southern end of a 9 mile (N/S) offensive and lies just north of the French town of Armentieres (although the wood is actually in Belgium). The Wood was referred to as Plugstreet by the British troops.
A British WWI machine-gun bunker deep in Ploegsteert Wood.
John’s brother William was in the 35th Australian Infantry Battalion, which fought in the Battle of Passchedaele. He was killed on 12 October, 1917 at Passchendaele, five months after the death of his brother. In a letter dated 21 October 1918, a Private J. Morris reported:
“He stayed behind a pill box at Passchendaele and said he was going to get the German gun to use it against them. He never came back to us. A Pte. Fred Walker reported that Sergt. Curtain told him in December that at Passchendaele on Oct. 12 1917, Melrose was killed instantly by a shell.”
Will Melrose is also remembered on the Menin Gate, mid right column,Wild Teasel in Ploegsteert Wood
Just over 110 years ago my grandfather William Melrose Mathers (1893-1953) and his brother Thomas (1891-1963), enlisted in the Australian Army on 2 September 1915. Being strongly opposed to killing anyone, they managed to get assigned to the 8th Field Ambulance of the Australian Medical Corps. They spent the next three and a half years on the Western Front in France and Belgium, on the front lines and at Regimental Aid Posts (RAPs) and hospitals behind the front lines.
Will Mathers (left) Will and Tom in Salisbury, 2 September, 1916
My older son went on a school trip to the Western Front battlefields in 2018 and visited battlefields where his great-grandfather had served as part of the Australian Field Ambulance. My younger son and his girlfriend both missed out on similar trips in 2021 due to the covid restrictions. So we went on a week-long trip to Flanders and the Somme to see some of the important sites on the Western Front. Our first stop was the Flemish city of Ypres and the battlefields nearby.
The medieval city of Ypres was completely leveled by artillery fire between 1914 and 1917. The city’s 13th-century Cloth Hall and St Martin’s Cathedral, seen in this photo, were reduced to rubble.
Ypres: the 13th-century Cloth Hall and St Martin’s Cathedral in 1917
After the war, some wanted to leave the city untouched as a reminder of the destruction, but the Flemish residents decided to rebuild the city faithfully reproducing the medieval buildings, including the magnificent Cloth Hall, in this photo, which houses the Museum “In Flanders Fields”.
The rebulit Cloth Hall todayThis heavy 42cm artillery shell stands as a stark monument to the relentless bombardment the city endured. Shells like this were fired from a “Big Bertha” siege howitzer.View of the rebuilt St. Martin’s Cathedral from the Belfry Tower of the Cloth HallLooking towards the Menin Gate.
The Menin Gate is dedicated to the British and Commonwealth soldiers who were killed in the Ypres Salient of World War I and whose graves are unknown. The names are recorded on stone panels of 54,395 Commonwealth soldiers who died in the Salient but whose bodies have never been identified or found. These include the names of 6,198 Australians. They also include the names of the brothers John and William Melrose, Scottish cousins of my grandfather William Melrose Mathers. Every night at 8 pm, the Last Post is played at the Menin Gate.
Crowd gathering at the Menin Gate for the Last Post
The Third Battle of Ypres, also known as the Battle of Paschendaele began on 31 July 1917. Massive artillery fire on the eve of the battle was intended to destroy German positions, but failed to do so. By mid-August, the first day objectives had still not been reached. A month later, the offensive resumed. New Zealanders, Australians and South Africans replaced British trooped in the spearhead of the attack.
The 8th Field Ambulance served on the Somme until September 1917, when it moved to Ypres to support the Australians. On 20 September, Will and Tom went into the front line for 48 hours to bring out the wounded on stretchers. They had a day in a rest area before going back into the front line on 23 September at Hooge.
Trenches at Hooge Crater were reinforced with corrugated iron. The pre-shaped, semi-circular corrugated iron sheets used by both Allied and German forces to construct and reinforce overhead coverings were known as “Baby Elephants”.
A trench at Hooge Crater
Hooge was the site of several large German mine craters. The lake in the photo below fills three German mine craters blown in June 1916 as part of their offensive against the Canadians.
Hooge Crater todayOne of many piles of artillery shells.A baby elephant is in the background.
One of many piles of artillery shells. In the two weeks before the attack alone, 4.2 million shells were fired at the German positions. The shelling failed to dent the German defences, but it destroyed the landscape and its drainage. The poor state of the terrain made it hard to move the guns forward.
A coil of barbed wire.
Will and Tom spent 72 hours evacuating the wounded from waterlogged trenches, dug outs and shell holes. Other members of the unit worked at the RAPs under fire. The bearers at Hooge endured heavy shell fire and the unit received many casualties.
The line of low hills running east of Ypres includes Hooge, Polygon Wood and Passchendaele. The Battle of Polygon Wood (26 September to 3 October 1917) took place in woodland that had been almost totally destroyed by the huge bombardments from both sides since 16 July and the area had changed hands several times. The I Anzac Corps (under Lieutenant-General William Birdwood) conducted the main advance of about 1,200 yd (1,100 m), to complete the occupation of Polygon Wood and the south end of Zonnebeke village.
Australian infantry attack on Polygon Wood. Painting by Fred Leist 1919. Me in a regrown Polygon Wood.This German bunker,built in 1916, was captured by the 56th Australian Battalion on 26 September 1917.Inside the bunker.
Will and Tom were gassed in Polygon Wood on 29 September 1917 and both became became sick. Will was hospitalized with gastritis for 8 days; Tom with a hernia of the groin was put on light duties.
Australian troops Australian infantry with small box respirator gas masks, Ypres, September 1917. The Buttes Cemetery in Polygon Wood contains the graves of 1295 British, 50 Canadian, 564 Australian, 163 New Zealand and 30 unidentified. Only 428 of these could be identified by name. Overlooking the cemetery is a monument to the 5th Australian Divison, which fought here.
After walking through Polygon Wood with temperatures around 32 C, we found ourselves at the Anzac Rest where a refreshing beer was welcome.
Australian hat collection on the ceiling beams.Felix uses a periscope in a trench at Paschendaele. Bunker and trench at Passchendaele.
As the Battle of Passchendaele continued, more focused attacks gradually shifted the front line but the losses were huge. And the artillery bogged down in the mud. In April 1918, to reinforce lines against the massive German Spring Offensive (Operation Georgette), the British evacuated the heavily contested Passchendaele Ridge, surrendering ground that had cost thousands of lives in 1917.
The allies were back where they had been in 1914. Their well developed positions had been exchanged for a pulverised, smaller bulge where everything had to be built afresh. That captured terrain had been transformed into a moonscape and the territory gained, at just 8 kilometres, came at a high price, with an estimated 600,000 casualties, including more than 125,000 dead. Passchendaele became an international symbol of the cruelty and futility of total war.
Tyne Cot War Cemetery, Passchendaele.
Tyne Cot is the Largest Commonwealth wall Groves cemetery in the world. There are almost 12,000 graves and a memorial wall bears the names of 35,000 soldiers who have no known grave. Most of those commemorated here died during the Battle of Passchendaele.
Tyne Cot Memorial Wall
In 1917, the site was completely barren except for German concrete bunkers with Machine gun nests. On 4th of October, the 40th battalion of the third Australian division broke through the position at Tyne cot. Over the following days, ANZAC troops tried in vain to advance from here towards Passchendaele. The bunkers on the site were converted to medical dressing station shortly after being taken. Those who succumbed to their injuries were buried at this site. This was the start of the cemetery.
In my earlier article, Questions about Panpsychism, I thought through what a defensible form of panpsychism might look like, and what questions it raises. I have now taken a dive into recent philosophical literature and debates on panpsychism. I focused particularly on Nagel’s 1979 article on panpsychism and three recent books on panpsychism which also include essays and commentaries by philosophers and scientists (Goff 2019, Goff and Moran 2022, Strawson 2024). I discovered that almost all of them ignore direct evidence and rely on assumptions and argument, never a good approach to understanding the nature of reality.
What philosophers and scientists think about panpsychism
In total, these four publications include essays and commentaries by forty-two authors. I am counting Goff (2006) and Goff (2019, 2022) as two separate authors, as his views changed substantially over this period. These 42 authors comprise 29 philosophers, 11 scientists (mainly physicists and neuroscientists), and two theologians.
Thomas Nagel (1979). Panpsychism.
Thomas Nagel’s 1979 article “Panpsychism” arguably started the recent resurgence of interest in panpsychism among philosophers. Nagel assumes panpsychism is about mental states, which he defines as “thoughts, feeling, emotion, sensation or desire”.
He argues that there are no truly emergent properties of complex systems. Nagel defines emergence as referring to an observed feature of a system that cannot be derived from the properties currently attributed to its constituents. In particular, subjective conscious experience cannot be derived from the purely objective physical properties of matter. Nagel argues that this means that mental states must go all the way down to the fundamental constituents of matter.
Rather than accept this conclusion, Nagel states that panpsychism must be added to the list of hopelessly unacceptable solutions to the mind-body problem. Why? Because it seems obviously ridiculous to ascribe proto-minds to atoms or electrons
In his concluding paragraph, Nagel identifies the so-called combination problem, which most of the philosophers in these books consider the biggest problem with panpsychism. He cannot imagine how mental states of animals could be explained in terms of the combination of the proto-mental properties of dead matter. He says “Presumably the components out of which a POV is constructed would not themselves have to have points of view. How could a single self be composed of many souls?”
Galen Strawson (2024) Consciousness and Its Place in Nature.
The second edition of Consciousness and Its Place in Nature includes Strawson’s original 2006 paper “Realistic Monism: Why Physicalism Entails Panpsychism” (Strawson 2006) and responses by 18 commentators, all philosophers apart from one physicist. Only two of the philosophers could be called panpyschists, though one of the two is generally critical of Strawson. The book also includes a lengthy reply by Strawson to the critiques from the 18 commentators, plus additional new commentaries by six of the original commentators.
Most philosophers and scientists assume that consciousness is an emergent property of brains. Strawson strongly rejects this. His argument is worth quoting as it is the key motivation for exploring panpsychist ideas.
“It is built into the heart of the notion of emergence that emergence cannot be brute in the sense of there being absolutely no reason in the nature of things why the emerging thing is as it is (so that it is unintelligible even to God). For any feature Y of anything that is correctly considered to be emergent from X, there must be something about X and X alone in virtue of which Y emerges, and which is sufficient for Y. ……. One problem is that brute emergence is by definition a miracle every time it occurs, for it is true by hypothesis that in brute emergence there is absolutely nothing about X, the emerged-from, in virtue of which Y, the emerger, emerges from it.” (Strawson 2024, p36)
Current science provides no examples of emergent phenomena that are brute, to use Strawson’s term. Emergent behaviours such as chaotic dynamics are qualitatively different but still explicable in terms of the underlying physical properties of its components. In what follows, I will use the term strong emergence to refer to this, and weak emergence to refer to emergent phenomena that are explicable in terms of underlying constituent properties.
Strawson argues that strong emergence of consciousness is essentially a magic passage across the divide between unaware matter and aware consciousness. Yet that magic passage is perfectly law-like every time it occurs. Humans start off as a single cell and yet all develop consciousness and subjective experience. And at least according to their reports, consciousness behaves in similar ways for most humans most of the time and behaves in predictable ways in response to various forms of meditation or methods for altering states of consciousness.
Despite this, most neuroscientists, physicists and philosophers assume that consciousness must be an emergent property of non -aware matter. Given the lack of any proposed mechanism for such strong emergence, this is similar to the situation with theist apologetics arguments which simply assume the universe must have had an uncreated creator who magically created it.
The 17 philosophers who comment on Strawson’s article include four who don’t think consciousness or experience exists and another nine who assume the word consciousness refers to minds (thoughts, feelings, experiences) and reject panpsychism as ridiculous (how could particles have thoughts or feelings) or because of the combination problem. The latter include Goff, who in 2006 thought the combination problem was fatal to panpsychism.
Philip Goff (2019) — Galileo’s Error: Foundations for a New Science of Consciousness
According to Goff, Galileo’s error was to exclude sensory experience from science in favour of what can be observed, and this has meant science cannot solve the hard problem. Goff wants to bring consciousness within the remit of science.
The 2019 Goff has moved some distance from the typical view of panpsychism expressed by the Strawson commentators (including the 2006 Goff) of electrons having thoughts and feelings, etc. He notes that “if electrons have experience, then it is of some unimaginably simple form”. And that complex conscious experiences such as humans have “is the result of millions of years of evolution by natural selection, and it is clear that nothing of this kind is had by elementary particles”. He also concludes that though the electrons in a sock may have a very elementary experience, that does not make socks conscious at the sock level.
But he doesn’t have any real suggestions about how to do this. He argues that:
“We must move to a post-Galilean paradigm, in which the data of consciousness and the data of physics are both taken seriously. Nothing less than a revolution is called for, and it’s already on its way.” (Goff 2019)
All fine as far as it goes. Except that the only data of consciousness he mentions is philosophical speculation. Although he mentions elsewhere that he meditations, there is no mention of the actual study of consciousness via meditation, psychedelics, or other forms of direct exploration. And why he focuses on the data of physics rather than neuroscience I have no idea.
Philip Goff and Alex Moran (2022). Is Consciousness Everywhere: Essays on Panpsychism
These essays first appeared as a special issue of the Journal of Consciousness Studies in 2021. The contributors included ten scientists, ten philosophers and two theologians. In the final article of the book, Goff responds to the essays in the volume and explores some ideas on what a “post-Galilean” science of consciousness might look like.
In 2006, Goff was negative about panpsychism because he thought the combination problem was unintelligible (thousands of small experiences coming together to be one big experience). He changed his mind in Strawson (2024) and talks about switching from a particle to field view of reality — without ever developing this.
Goff makes the point that consciousness is not a normal scientific phenomenon, publicly observable by everyone.
“In the unique case of consciousness, the thing we are trying to explain is not publicly observable. …. There is information that we get from attending to our experience, information that cannot be conveyed in the language of physical science.” (Goff 2022)
However, he then essentially dismisses information derived from systematic direct observation of consciousness, concluding that
“Because consciousness is not publicly observable, this is not a question one can answer with an experiment (all experiments can do is establish more correlations [with neural happenings). At this point we must turn to philosophy, examining the various proposals philosophers have offered to account for the fact that brain activity is correlated with experience.” (Goff 2022)
Unfortunately, ignoring the collective first-person experience of consciousness and relying on assumptions, arguments and speculation has led philosophers to a version of panpsychism that is inconsistent with the available evidence and rightly rejected as nonsensical by most of them and by many others who take an interest in consciousness. In what follows, I will identify important evidence from first-person experience that allows the development of a much more plausible form of panpsychism.
1. Taking first-person evidence into account
Goff [2019] notes that there is a fair amount of scepticism at the idea that philosophers might have anything to contribute to the scientific project of uncovering the nature of reality. He adds “Such scepticism is presumably rooted in the fact that, in general, philosophers reach their conclusions without actually performing any experiments or carrying out any observations. In contrast to the natural scientist, the principal activity of the philosopher is thought.”
Although he called for taking the data of consciousness into account, he remained stuck in philosophical speculation. Data of consciousness can only come from first-person observations, the actual systematic study of consciousness via meditation, psychedelics, breathwork, and other techniques for exploring consciousness and non-ordinary states of consciousness.
Thought experiments without a basis in observation or experiment are meaningless. Millenia of religious apologetics have demonstrated that. Largely missing from the debates among the philosophers and scientists is any consideration of the evidence from direct observation of consciousness. Christoph Koch realizes this in his commentary in Goff and Moran (2022):
“The last 2,500 years of Western philosophy of mind have seen the rise and fall (and rise and fall) of many schools of thought concerning the mental and the physical. Highly polished arguments and counterarguments are exchanged in a never-ending cycle that results in drawn-out sophisticated disagreements but no resolution.
“Indeed, it is not even clear that there is a generally accepted notion of progress within philosophy of mind (Bourget and Chalmers, 2014; Chalmers, 2015). …… The science of consciousness must break out of these endless epicycles of arguments by formulating a sustainable programme of hypothesis formulation and experimental validation or falsification. (Koch 2022)
Neuroscience is only one part of the needed inputs; it can explore the functioning of the brain and the objectively measurable correlates of conscious states; the other more important part has to come from systematic direct observation of consciousness. Currently, the two major approaches to systematic first-person exploration of consciousness are meditation and psychedelics.
Of the 42 philosophers, scientists and theologians included in the four books I read, only three (all philosophers) discussed meditation as a source of data on consciousness and only one (a theologian) discussed the potential use of psychedelics as a source of data. None of them attempted to use data from direct observations in their contributions.
2. Consciousness and contents of consciousness
Contents of consciousness include thoughts, feelings, brain-processed sensory inputs, etc. Neuroscience has provided convincing evidence that these contents are all produced in brains. Most beginning meditators will achieve this insight relatively early in their practice, as they see content come and go in the field of awareness untouched by an attempt at conscious control. It often takes more sustained practice to achieve a state where brain content dies away completely for some length of time, leaving a pure experience of contentless awareness (see my previous article).
This contentless awareness (also referred to as formless or empty) is what I define consciousness as referring to. Until reading these books, I had not fully grasped that most of the writers on panpsychism assumed that consciousness referred to minds, or selves, or the full contents of consciousness. In fairness to me, I must also point out that hardly any of the philosophers attempt to specify what they are using the word consciousness to refer to, beyond a vague invocation of experiences or qualia.
The primary motivation for panpsychism is the strong emergence problem, and clearly (1) contents of consciousness are of course products of (weak) emergent brain processes and (2) to solve the strong emergence problem, consciousness (ie. formless awareness) is all that needs to go “all the way” down.
Of the 42 philosophers, scientists and theologians, only 3, maybe 4, philosophers (Strawson, ) are some ways towards this insight. Of the rest, apart from several who deny consciousness exists, all imagine the word refers to minds with thoughts, feelings, experiences, and that these are what go all the way down. Just to quote a few examples:
“Will each type of particle possess a wide range of experiences, including emotions and thoughts, or will particles specialize in certain types of experience — electrons doing sensory, protons handling the emotional, neutrinos taking care of the cognitive?” — McGinn (2024)
“The idea of electrons making decisions about how to spin, nuclei harbouring intentions to split, or photons with existential Angst, makes idealism seem positively sane.” (Simons 2024)
“In what way could such a mental aggregate [anticipating a cold beer] consist of or be determined by or otherwise ‘arise from’ a swarm of smaller mentations? Is it that some of my ultimate components are experiencing some of those very same mental states, and when enough of them do, I myself do?” (Lycan 2024)
Strawson himself clarifies that “by panpsychism I mean the view that the basic physical constituents of the universe have mental [and in particular experiential] properties” (Strawson 2024a, p. 318). He explicitly states that he uses the terms experience, experientiality, conscious experience, consciousness interchangeably. He considers experiences of colour, taste, smell, pain, fear, and so on as paradigm cases of experience, but conscious thought is also a case of experience.
A few of the philosophers come close to realizing that consciousness and its contents are not the same thing. Coleman does make a distinction between mind and consciousness, but then continues to use consciousness as a synonym for mind.
“Panpsychism, if we are guided by etymology, has it that mind is everywhere. But by contemporary usage it is, more strictly, the claim that consciousness is everywhere, and that it is fundamental.” Colman (2024)
Seager (2024) also struggles to articulate an insight that mind does not have to be what goes all the way down but does not have either the direct experience or the vocabulary to articulate it. He talks about a more primitive notion of “presence” being what is ubiquitous:
“Presence can be without being presence to mind. Perhaps it could be argued that ‘mind’ should be taken in some ultra minimal sense and that therefore presence resolves into infinitesimal sparks of consciousness. Perhaps this is only a verbal dispute, but such sparks are not what one would call conscious minds.” (Seager 2024)
Goff (2019, p. 206) mentions formless consciousness, a non-dual state of consciousness that mystics claim “is the backdrop to all individual conscious experiences”. But he has not experienced it himself and appears to have no understanding of how it solves all his issues with panpsychism. He thinks of formless consciousness as a supernatural state, the ground of being, and does not realize that basically it is just contentless awareness.
Realizing that only consciousness (contentless awareness) needs to go all the way down to avoid the strong emergence problem avoids the implausibility of electrons have minds, experiences, thoughts, selves and also solves the combination problem (see Section 5).
3. Consciousness and no-self
It is plain to most philosophers that there cannot possibly be experience — experiencing, — without a subject of experience (Strawson 2024b). Even Goff (2019) rejects the idea that he could experience different forms of self, or no-self: “Similarly, I cannot fully know what it’s like to be a split-brain patient, as I cannot adopt the perspective of someone whose consciousness is fragmented into isolated pockets.”
Again, we see the extreme limitations of philosophy. He has no experience of transforming his sense of self so simply, but confidently, just assumes it cannot be done. I have had a psychedelic journey in which I experienced several profound transformations of my sense of self, including a period where there were four selves. And several times while meditating, I have experienced body and mind dropping away, leaving simply contentless consciousness without any sense of self.
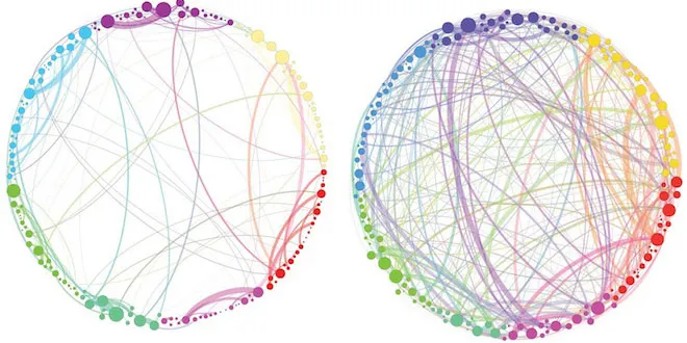
What do we mean by no-self or that the self is an illusion? Here the data from meditation and from neuroscience together give us a coherent explanation. Many experienced meditators report experiencing the cessation of any sense of self, leaving simply an awareness which may or may not have content. Neuroscientists have identified the sense of self as arising in the default mode network (DMN) which is responsible for autobiographical memory and ruminations (Davey and Harrison 2018, Seth 2021, Menon 2023, Azarius 2025). Functional magnetic resonance imaging of the brain has shown that psychedelics downregulate and modify the connectivity of the DNM, likely explaining how psychedelics can modify or eliminate the sense of self ((Carhart-Harris et al. 2014, Gattuso et al 2023, Benes et al 2025).
Communication between brain networks in people given psilocybin (right) or placebo (left). (Petri et al 2014)
Buddhists and many other meditators refer to the self as an illusion. This is somewhat misleading. The sense of self is real content experienced in everyday consciousness. That it is produced by the DNM means that it is a product of the brain in the same sense that a dream or hallucination is. It can change or disappear and is an illusion in the sense that it is not a fundamental and permanent essence of a human being. We can obviously conclude that the “many little selves” many philosophers agonize about, how they can combine to form a single big self, is all irrelevant.
Anneka Harris (2022) identifies the illusion of the self as the solution to the problem of particles having minds and of the combination problem. I would argue that she misses a much simpler conclusion. The “self” is only one of the contents of consciousness, and there is no need for panpsychism to be saddled with any of the other contents of consciousness going all the way down. It is much simpler to identify contentless awareness as all that needs to go all the way down to the basic constituents of everything.
Harris gets very close to this realization when she distinguishes between consciousness and thought. And says we would not expect a rock to have a single unified point of view (as a rock) or to have anything like thoughts or intentions. Because the rock particles are not configured to do that. She is so close to recognizing that brains produces all the contents of consciousness.
4. The supposed combination problem
Goff (2019, pp.144ff) describes the combination problem “how little minds could somehow combine to make up a big mind” as the biggest problem for panpsychism. Without a solution, panpsychism is a lost cause. Most of the 42 experts also see the combination problem as the biggest problem facing panpsychism.
The problem just goes away once we realize that only consciousness (defined as contentless awareness) needs to go all the way down to solve the hard problem without invoking strong emergence. I think there is also good evidence from direct experience to suggest that contentless pure awareness must be identical everywhere, for essentially the same reason that electrons are perfectly identical. There are no properties of electrons that differ across electrons. There is no property of contentless awareness per se that can differ across instances though any contents can and will differ.
Why does the combination problem not exist? Consider atoms, or their fundamental particle constituents, which have mass and hence a small gravitational field. There is no combination problem for larger aggregations of atoms. Whether it is a cannonball or a planet, there is a single gravitational field resulting from the combined mass of the constituents. Why should contentless awareness be any different? Following this analogy, the only aspect of contentless awareness that might vary across instances of it is the “strength” of the awareness “field”.
However, I think there is a more useful physical analogy for understanding how contentless awareness might combine across the constituents of a brain and interact with content produced by that brain. Let us consider a bar magnet. Individual atoms have magnetic fields arising from electron spin and motion and align within a microscopic domains to create an overall magnetic field. These domains are unaligned in an unmagnetized metal bar.
When they do become aligned (say by a strong magnetic field from an external source), then the overall metal bar becomes a permanent magnet with a north and south pole and a net magnetic field running between those two poles outside the bar. Normally, most of the domains are aligned though increasing temperature will result in fewer domains being aligned, and beyond a specific temperature, alignment ceases and the bar is no longer magnetized.
Perhaps consciousness of atoms or molecules combines in a similar way to form consciousness domains, and these combine to produce a single overall consciousness in a normal brain. What is combining is pure contentless awareness, which is not separate any more than the magnetic fields of the domains remain separate in a permanent bar magnet. That awareness experiences contents of contents produce by the related brain.
For simplicity, I will refer to the smallest amount of consciousness (say at electron or hadron level) as a quantum of consciousness. I am not invoking quantum theory here, in any way, just borrowing a piece of terminology.
Why are conscious subjects at the human scale separate in terms of the experiences in consciousness? Biologists tend to see this as an argument for emergence from brains. The real answer is likely in what is needed for quanta of consciousness to form a broader field at organ or organism level in such a way as to have causal connections with brain products.
5. The evolution of conscious minds
As nervous systems and brains evolve, their capacity to create and process thoughts, feelings, sense experiences generally increase. If the basic constituents of these nervous systems have a primitive contentless awareness, which can combine into a single larger field of awareness, how and why do the contents of awareness appear in that field? In what follows, I will refer to brains for simplicity but intend that reference to cover “brains and associated nervous systems”.
Is this a causal process from brain to consciousness or is the “eye of awareness” just aware of content without causal connections. When I read Chalmers (1996), he semi-convinced me that pure consciousness was an epiphenomenon that did not causally affect its contents or the neurons producing them. There is no doubt that those contents, produced by the brain, are experienced in consciousness. Does that experiencing in turn have causal effects on the brain?
While writing this article, I have been examining my meditation experience for any clues. I do have a memory of what my most profoundly non-dual state of consciousness was like. In fact, it made a huge impression and was a major reason why I spent the next decade practicing zen meditation in week-long retreats under the guidance of several Zen master.
I tentatively conclude that memories of largely contentless states of consciousness do get made. Presumably by a causal pathway from consciousness to memory-making processes of the brain. And it follows that the experiencing in consciousness of content produced by the brain also has a causal feedback mechanism to the brain.
Why is it useful for humans to “experience” with feedback to the brain, on top of collecting and processing sensory and other data? Or put slightly differently, why would evolution select for brain structures that supported increasingly strong causal connections with consciousness?
“Among all the aspects of an animal or human being that contribute to its fitness, few make a greater contribution than consciousness. There is much evidence that the focus of a person’s consciousness can be trained and that a trained attention underlies the skills of an athlete, a musician, or a hunter. It seems possible, if not likely, that consciousness or awareness had and has a lot to do with the thriving of our species.
“It is then very natural to suppose that, if the existence of consciousness is to be explicable, it must perform some function that increases the fitness of the creature that is endowed with it. But this requires that consciousness can intervene in the network of causes in the physical universe. But when we try to develop this idea, we run immediately into a very strong argument that the physicalists have to their credit, based on the causal completeness — or ‘causal closure’ — of the of the standard Newtonian paradigm” (Cortês, Smolin and Verde (2022).
Cortês et al. conclude that there must be a yet unknown causal pathway between the experience of qualia and the brain. It makes the will to live and to reproduce more real and motivating to the organism. An unconscious brain can be programmed with these drives, but there is likely some additional oomph provided by the “experiencing” of things.
The psychologist Nicholas Humphrey (2017), who thinks consciousness is an illusion, has argued that it comes with a significant survival advantage. Creatures who believe they have a subjective inner life develop a great interest in preserving and enriching that inner life through complex engagement with their environment.
Strawson (2006) also realizes that natural selection will act on experiential consciousness:
“Human experience or sea snail experience (if any) is an emergent property of structures of ultimates whose individual experientiality no more resembles human or sea snail experientiality than an electron resembles a molecule, a neuron, a brain, or a human being. Once upon a time there was relatively unorganized matter, with both experiential and non-experiential fundamental features. It organized into increasingly complex forms, both experiential and non-experiential, by many processes including evolution by natural selection. And just as there was spectacular enlargement and fine-tuning of non-experiential forms (the bodies of living things), so too there was spectacular enlargement and fine-tuning of experiential forms.” (Strawson 2024a, p48)
However, because he thinks it is experiences which go all the way down to fundamental constituents, he misses what natural selection is really doing. As primitive nervous systems evolved into more complex organization of neurons and neurotransmitters, natural selection will give an evolutionary advantage to forms of organization which align basic quanta of consciousness at molecular level into a coherent field of consciousness that can have experiences and give causal feedback to the nervous system.
What natural selection must have done is evolve the alignment of basic quanta of consciousness into a larger field of consciousness in such a way as to have two-way causal paths between that field and the associated brain. Natural selection will also be fine-tuning brains to produce more complex contents to be experienced in consciousness.
Complex brain content is weakly emergent, and we already largely understand how thought, feelings and sense experiences arise in terms of the properties of the basic constituents of brains. Contentless organism-level consciousness is also now weakly emergent since its properties are entirely explicable in terms of the quanta of contentless consciousness of the basic constituents of matter.
6. Towards a defensible form of panpsychism
The panpsychism theory proposed in this article can be summarized as follow:
Consciousness is real, it exists.
The term consciousness refers to contentless awareness per se, not to mind or self or thoughts or qualia or other contents of consciousness.
Consciousness cannot be strongly emergent from purely physical constituents.
Some degree of consciousness is a property of the basic constituents of matter.
Contents of consciousness (thoughts, feelings, sense experiences) are produced in brains and experienced in consciousness.
Without the involvement of a brain (or perhaps nervous system), there are no thoughts, no thinking, no feelings etc just some degree of contentless awareness.
There is no combination problem for basic quanta of consciousness.
Either such quanta naturally form a single field of contentless awareness in aggregates of matter, or the alignment and combination of elementary quanta of consciousness only occurs in specific circumstances (e.g., in brains evolved to provide those circumstances)
Consciousness connected causally with brain content provides an evolutionary advantage and (soft) emerges in organisms under the influence of natural selection.
Contents of consciousness are emergent phenomena, a soft emergence associated with the evolution of increasingly complex nervous systems and brains.
Given the long use of the term “panpsychism” for proposals based on a mind-body dichotomy rightly rejected by most philosophers and lay people as clearly nonsensical, we need a distinguishing label for this theory. I will provisionally call it emergent panpsychism, since both consciousness and brain contents are weakly emergent and there is no longer a hard problem of strong emergence.
Some of the commentators criticized panpsychism on the grounds that it changes nothing and has no testable hypotheses. It will be important to try to clarify the evidence for causal connections between consciousness and brain processes. This will necessarily involve systematic exploration using techniques such as meditation.
Luke Roelofs (2022) notes that the claim that panpsychism and a theory of hard emergent consciousness are observationally equivalent has been used by critics to argue that panpsychism is unfalsifiable. Roelofs argues that they do differ in their predictions of whether or not I am subjectively conscious: panpsychism predicts that I should be, while materialism gives no basis for predicting this.
Emergent panpsychism likely has a number of other predictions. I have not thought about this much yet, but will list a few predictions off the top of my head:
Consciousness, qualia, and a sense of self are not confined to humans, but present in other animals and organisms to varying degrees (likely correlated with the complexity of the nervous system).
The emergence of consciousness and causal connections to brains is due to natural selection. There is negligible probability that the relevant structures and interactions required for consciousness will occur in computer circuits built by humans entirely ignorant of them.
Until and unless we discover such knowledge, digital processes and AI programs will never become conscious. Not even genuine “thinking” AI of the future, let alone the massive statistical autocomplete software we currently call AI.
Contentless awareness is likely a single universal field, but content is only ever experienced locally in relation to specific organisms. Content cannot be shared across organisms or shared communally except through linkages between brains.
Strawson made some quite similar predictions in a recent interview (Andrew 2025):
“Yeah, I would think that of larger things that are conscious, probably they’re all biologically evolved. That would be my bet. Well, now it’s partly because I think that interesting animal consciousness biologically evolved for a purpose, and that wouldn’t happen in the case of the chair.
“And, empirically, I think it’s highly plausible that you need some incredibly complicated electrochemical shenanigans to get interesting consciousness — and chairs don’t have it. My brain has it, and the molecular structure of a chair just doesn’t have the kind of electrochemical goings-on of the sort that would be needed for interesting consciousness.”
To be clear, Strawson here is talking about the emergence of interesting animal consciousness from existing basic quanta of consciousness, not about hard emergence from purely physical materials.
8. Conclusions
Having done a deep dive into philosophers’ debates on panpsychism, I was very surprised to find that almost all of them think panpsychism refers to minds, even little selves, and then they get stuck in the combination problem “how can lots of little minds combine into one big mind?” More importantly, this erroneous dualism leaves the contents of consciousness (thoughts, feelings, experiences) on the side of mind, and even most philosophers instinctively recoil from a version of panpsychism in which spoons and toasters have minds, and hence thoughts, feelings and experiences. This is why Strawson rejected panpsychism after writing an article about the need for it.
The books reviewed here included considerable discussion of Descartes, the mind-body dichotomy and the problems of dualism. A committed practice of meditation, even for a few months, will usually yield the insight that contents of consciousness come and go, in a field of contentless consciousness. Once that distinction is made, it becomes obvious that strong emergence only requires pure contentless consciousness to go all the way down — leading to the emergent panpsychism I have outlined in this article.
Even those philosophers like Chalmers, who seems passionate to understand the nature of consciousness, ignore systematic observations. In a 2017 interview with Chalmers, John Horgan (2017) reported that Chalmers has “never had the patience” for meditation, and he has doubts about basic Buddhist claims, such as anatta, the insight that the self does not really exist. What would we think of a physicist who never had the patience for experiments, said he also doubted the evidence of the experimentalists (or never actually looked at it) and preferred to do thought experiments based on assumptions that seemed reasonable to him?
This is main reason that there has been no real progress in the philosophy of mind or consciousness over many centuries (see Koch 2022, quoted in Section 3 above). So why do even the philosophers passionate about the nature of consciousness ignore direct first-person evidence? Anyone serious about elucidating the nature of an aspect of reality would surely be willing to devote years of research to either the systematic first-person exploration of consciousness or the collective reporting of those who have done so. Are these philosophers simply not serious?
I think there are several reasons for this. First, they may be stuck in a cultural bubble which includes the mind-body dualism paradigm and a view of evidence restricted to the third-person data of science. Second, some may simply be playing the academic game of publish or perish and unwilling to risk going outside accepted academic disciplinary boundaries. Thirdly, some may see the major forms of direct evidence from meditation or psychedelics as unacceptable woo-woo or meaningless.
Strawson, Goff and a couple of other philosophers have come close to an understanding that only the simplest form of consciousness has to go all the way down but lack the direct experience of contentless consciousness that would allow them to articulate this clearly. Of course, no-one should just take a single person’s report or interpretation of their experience as gospel. Ideally you experience it for yourself, taking into account the advice of those who have traveled this direct experiential path, testing it, evaluating it for yourself. Persevere until you have experience and understanding of the main techniques, plus experiences of the various important states of consciousness that can be reached. Then you can join the conversation.
This article was earlier published on Medium here.
References
Andrew JP (2025). My Interview With Galen Strawson, Part I: On Galen’s background, his conception of philosophy, and panpsychism. Substack, 3 May 2025.
Azarias FR, Almeida GHDR, de Melo LF, Rici REG, Maria DA. The Journey of the Default Mode Network: Development, Function, and Impact on Mental Health. Biology. 2025; 14(4):395. https://doi.org/10.3390/biology14040395
Chalmers, David (1996), The Conscious Mind: In Search of a Fundamental Theory, New York: Oxford University Press.
Bourget, D. & Chalmers, D.J. (2014) What do philosophers believe? Philosophical Studies, 170, pp. 465–500.
Chalmers, D.J. (2015) Why isn’t there more progress in philosophy? Philosophy, 90, pp. 3–31.
Chalmers, David, and McQueen, Kelvin (2022). “Consciousness and the Collapse of the Wave Function,” in S. Gao, ed., Quantum Mechanics and Consciousness. New York, 2022; online edn, Oxford Academic, 20 Oct. 2022, https://doi.org/10.1093/oso/9780197501665.001.0001
Colman, Sam (2024). A Panqualityist Manifesto, In Goff, Philip and Moran, Alex (eds.). Is Consciousness Everywhere: Essays on Panpsychism. Andrews UK Limited 2022, Kindle Edition, p. 466-488.
Cortês, Marina, Smolin, Lee, and Verde, Clelia (2022). Physics, Time, and Qualia. In Goff, Philip and Moran, Alex (eds.). Is Consciousness Everywhere: Essays on Panpsychism. Andrews UK Limited 2022, Kindle Edition, p. 54-75.
Davey CG, Harrison BJ. The brain’s center of gravity: how the default mode network helps us to understand the self. World Psychiatry. 2018 Oct;17(3):278-279. https://doi.org/10.1002/wps.20553
Gattuso JJ, Perkins D, Ruffell S, Lawrence AJ, Hoyer D, Jacobson LH, Timmermann C, Castle D, Rossell SL, Downey LA, Pagni BA, Galvão-Coelho NL, Nutt D, Sarris J (2023). Default Mode Network Modulation by Psychedelics: A Systematic Review. Int J Neuropsychopharmacol. 2023 Mar 22;26(3):155-188. doi: 10.1093/ijnp/pyac074.
Goff, Philip (2006) Experiences Don’t Sum. In Strawson (2024). Consciousness and Its Place in Nature. Andrews UK Limited. Kindle Edition, p. 97.
Goff, Philip (2019). Galileo’s Error: Foundations for a New Science of Consciousness. Ebury Publishing.
Goff, Philip (2022). Putting Consciousness First: Replies to Critics. In: Goff, Philip and Moran, Alex (eds.). Is Consciousness Everywhere: Essays on Panpsychism. Andrews UK Limited 2022, Kindle Edition, p. 409-461.
Goff, Philip and Moran, Alex (eds.) (2022). Is Consciousness Everywhere: Essays on Panpsychism. Imprint Academic. Kindle Edition.
Harris, Annaka (2019). Conscious: A Brief Guide to the Fundamental Mystery of the Mind, New York: Harper Collins.
Harris, Annaka (2022). A Solution to the Combination Problem and the Future of Panpsychism. In Goff, Philip and Moran, Alex (eds.). Is Consciousness Everywhere: Essays on Panpsychism. Andrews UK Limited 2022, Kindle Edition, p. 187-202.
Koch, Christof (2022). Reflections of a Natural Scientist on Panpsychism, In Goff, Philip and Moran, Alex (eds.). Is Consciousness Everywhere: Essays on Panpsychism. Andrews UK Limited 2022, Kindle Edition, p. 94.
Mc Ginn, Colin (2024). Hard Questions: comments on Galen Strawson. In Strawson, Galen. Consciousness and Its Place in Nature: Why Physicalism Entails Panpsychism. 2nd Edition 2024, p.160-175.
Nagel, Thomas (1979), ‘Panpsychism’, in Mortal Questions. Cambridge: Cambridge University Press: 181–95.
Petri G, Expert P, Turkheimer F, Carhart-Harris R, Nutt D, Hellyer PJ, Vaccarino F (2014). Homological scaffolds of brain functional networks. J R Soc Interface 6 December 2014; 11 (101): 20140873. https://doi.org/10.1098/rsif.2014.0873
Roelofs, Luke (2022) Is Panpsychism at Odds with Science? In Goff, Philip and Moran, Alex (eds.). Is Consciousness Everywhere: Essays on Panpsychism. Andrews UK Limited 2022, Kindle Edition, p. 171-185.
Seager, William (2024). From Panpsychism to Neutral Monism… and Back Again(?). In Strawson, Galen. Consciousness and Its Place in Nature: Why Physicalism Entails Panpsychism. 2nd Edition 2024, p.509-532.
Simons, Peter (2024). The Seeds of Experience. In Strawson, Galen. Consciousness and Its Place in Nature: Why Physicalism Entails Panpsychism. 2nd Edition 2024, p.251-258.
Strawson, Galen (2006). Realistic monism – why physicalism entails panpsychism. Journal of Consciousness Studies 13 (10-11):3-31. Reprinted in Strawson (2024).
Strawson, Galen (2024a) Consciousness and Its Place in Nature: Why Physicalism Entails Panpsychism. 2nd Edition 2024.
Strawson, Galen (2024b) The Impossibility of Subjectless Experience. Journal of Consciousness Studies, Volume 31, Numbers 5-6, June 2024, pp. 26-36(11).
In the last two weeks of July 2025, I took my younger son to central Australia to visit for his first the iconic desert landscape and to explore some of the spectacular desert and mountain scenery. I have visited central Australia a number of times when I was younger, with trips to Alice Springs and Uluru (Ayers Rock), Katherine Gorge and the Arnhem Land Escarpment.
I have included links to previous posts about this trip and the end of this article. From Kings Canyon, we drove 330 km southwest to Uluru, formerly known as Ayers Rock, and perhaps the most famous landmark in Australia. As we headed towards Uluru, the land became extremely flat. The recent rain from the storm during the last night we were in Kings Canyon was widespread and had filled some of the shallow lakes that usually form only in the wet season.
One of the shallow lakes we saw on the way to Uluru, an unusual sight in the dry season.Mount Conner about 90 km before Uluru — aka Mount Fuluru because many people mistake it for Uluru.
We stayed at a tourist hotel in Yulara, a new settlement about 20 km from Uluru, built specifically to provide tourist accommodation and services, including an airport.
Approaching Uluru from Yulara.
This was my second visit to Uluru. The first was 45 years ago, one week before the dingo took Azaria in August 1980. That was also a memorable trip. One of my friends had found a pilot seeking to increase his flying hours, who was willing to fly the four of us from Sydney to Uluru in a light plane, all for the cost of the fuel. I think we paid a couple of hundred dollars each.
On that first trip, we slept in a cave at the base of Uluru, and the pilot slept under the wing of the aircraft. In those days, it was still possible to climb Uluru, and I climbed it before dawn the next day, to arrive at the summit just before sunrise and to see the shadow of Uluru stretching all the way to the horizon when the sun rose. My three friends were all rock climbers and had decided to do a free solo of Uluru on its other side. I waited on top till they arrived quite some time later, and they looked quite traumatized. It seems the gully they had chosen to climb ended on a steep face which was extremely exposed and pushed my friends well out of their comfort zone.
A brief anecdote. When first writing some notes about my first trip to Ayers Rock, I could not remember exactly which year the trip was, but I knew we were there the weekend before the dingo took the baby Azaria Chamberlain. The baby’s mother Lindy Chamberlain was convicted of murdering the baby, and spent three years in prison before the baby’s clothing was found in a dingo den, and she was exonerated. So I looked up this event on Wikipedia to check the date it occurred, and read the article. In the last paragraph it reported that Lindy and Michael Chamberlain had divorced after these event, Lindy remarried an American who lived in Australia, and the dingo changed its name to Sally and was adopted by a family in Florida. I went back to the article a few days later to get a screenshot for posterity, but unfortunately and eagle-eyed editor had removed the information on the dingo.
I am fairly sure that this cave at the base of Uluru is the one we slept in on my first trip to Uluru.Aboriginal art on the wall of the cave.My son and I did a complete circuit around the base of Uluru, an 11 km walk.
Our last night in Kings Canyon, there was a violent storm with strong wind and heavy rain. As a result there were waterfalls all over the Rock, apparently a rare sight in the dry season.
Waterfalls on the southern flanks of Uluru.
Climbing the Rock was banned in 2019. The track to the summit is still visible as a white line up the ridge that reaches the summit immediately above Felix. Back in 1980, I climbed Uluru via this route.The route was extremely steep towards the top, and there was a chain railing that climbers could use to prevent themselves slipping and falling. At least 37 people died while attempting to climb Uluru (Ayers Rock) since record-keeping began in the 1950s.
Westen end of Uluru, with the former climbing track visible as a white line of the ridge to the left of centre.
As we headed back to Yulara, the sun was getting low in the sky and turning the Rock red.
Late afternoon sun on Uluru.
The next day we went out to Kata Tjuṯa (the Olgas), 56 km west of Uluru. This was my first land visit to them. On my first trip in 1980, the pilot had flown us out to the Olgas and we flew low over them getting superb views. I’m quite comfortable with the Aboriginal name for Ayers Rock, but I must admit I have trouble getting Kata Tjuta into my memory banks, and still think of these iconic rocks as the Olgas.
Kata Tjuṯa (the Olgas)/Looking back to Uluru from the Olgas.Flowing water in Walpa Gorge, with Mount Olga on the right.Walking towards the Valley of the WindsView towards the Valley of the WindsLooking back as we leave the Olgas to head back to Yulara
Seeing as we could not climb Uluru, we decided to do another short helicopter flight over it.
Heading towards Uluru in a small helicopter
The next day we drove from Uluru to Darwin, a 500 km trip. And spent two days in Darwin, a fascinating city well worth visiting. In the 1990s, I had been collaborating with the Northern Territory Health Department on a joint project and visited Darwin a number of times, so already was quite familiar with it. However, these two days we were able to delve into the history of Darwin, and particularly its experience of being attacked by the Japanese during World War II. Perhaps I will do another post on that sometime.
Here are links to earlier posts on our explorations of central Australia in 2025.
Kings Canyon is located about halfway between Alice Springs and Uluru, by road its about 470 km from Alice. The canyon has been cut 100 m deep into the layered sandstone and shale over a period of 400 million years, creating one the most spectacular landscapes in central Australia. We climbed 500 steps to the western canyon rim, then walked through the dome-like sandstone formations of the “Lost City”, crossing over to the other side of the canyon at a waterhole known as the Garden of Eden, before returning along the eastern rim of the canyon and back down to the bottom of the gorge.
Looking towards Kings CanyonNearing the northern rim of the CanyonFelix and ColinIn the Lost CityThese ripples in the sandstone were formed in the shallow waters of a long extinct sea 400 million years ago.Felix in the Lost CityStarting the descent into the canyon to the “Garden of Eden”Garden of EdenStairs ascend to the southern rim of the canyon from the Garden of EdenOn the southern rimLooking across the canyon to the Lost CityThe start of the descent back to Kings Creek
We descended to the Kings Creek level and drove back to Kings Creek Station for the night, before heading towards Uluru (Ayers Rock) the next day.
Emus near Kings Creek StationThere were also several camels grazing near the stationThere was no light pollution in the desert at night and the stars were spectacular. Felix took this photo of the Milky Way with his iphone, handheld with a 10 second exposure.
When we returned to Switzerland, I told Felix we needed to watch The Adventures of Priscilla Queen of the Desert, a great 1994 Australian movie in which two drag queens and a transgender woman travel through outback Australia and visit some of the places we had just seen. So we both watched it and realized that the climactic scene where drag queens Tick, Felicia, and Bernadette climb to the a cliff and gaze across the landscape was filmed at Kings Canyon and we recognized the path they climbed to the top of Kings Canyon. We even walked through the narrow chasm in the photo below from the movie. Its now known as “Priscilla’s Crack.”
The drag queens in Priscilla’s Crack 31 years ago. The lead character in red is played by Hugo Weaving, who went on the play Elrond in The Lord of the Rings movies.
In the penultimate scene of the film, the trio stand triumphant on top of Kings Canyon’s south wall; they’ve survived not only the hike, but the existential threat of bringing Sydney’s drag culture to the outback. The grandeur of the 400-million-year-old landscape in the sweeping panorama only magnifies the accomplishment. “It never ends, does it? All that space,” Terence Stamp’s character, Bernadette, remarks.
After visiting the West McDonnell Ranges (see previous post), we headed for Kings Canyon, which I had not visited before. Kings Canyon is located about halfway between Alice Springs and Uluru, by road its about 470 km from Alice. The canyon has been cut 100 m deep into the layered sandstone and shale over a period of 400 million years, creating one the most spectacular landscapes in central Australia.
We had booked a tent for two nights at Kings Creek Station, about 36 km before Kings Canyon. When we got there, we discovered that they had a small helicopter, and arranged to take a flight over Kings Canyon. This was Felix’s first time in a helicopter, and it was the smallest I had ever been in. The pilot took the doors off it so we could have unobstructed views.
Below is a short video and a few photos from the air. The views were spectacular. It was extremely windy in the open helicopter as we were flying at 180 km/hr and there were strong cross-winds. It was quite exhilarating to be flying with the wind buffeting us and essentially just a framework of metal around us open to the sky around us and the ground below.
My younger son Felix and I visited relatives in Australia in July last year. In the last week of our trip, I took Felix to the “red centre” of Australia to explore some of the spectacular desert and mountain scenery. I have visited central Australia a number of times when I was younger, with trips to Alice Springs and Uluru (Ayers Rock), Katherine Gorge and the Arnhem Land Escarpment. Felix is an Australian by descent, but had never visited the centre before.
We flew from Brisbane to Alice Springs to spend ten days in the Northern Territory. We hired a four-wheel drive car for a week, and spent our first day exploring Alice Springs. We then spent a day in the West MacDonnell Ranges, driving 140 km west of Alice Springs and visiting some of the many spectacular gaps and gorges as well as areas of Aboriginal significance.
Simpson’s Gap, 24km from Alice Springs. The rock walls are home to the rare and endangered Black-footed Rock Wallaby. Felix managed to spot one when he was scrambling around.Standley Chasn is a 25 minute walk along a natural creek bed from the road head, through acaciasm cycads and ghost gums, until the dramatic 80m sheer rockfaces of the Chasm are reached.MacDonnell Ranges in the distanceThe Dryland Feather-Head (Ptilotus xerophilus) was only recognized as a separate species restricted to central and western Australian in 2019, distinct from a similar eastern Australian species.Although they rise only 400-500 metres from the surrounding mulga scrub, the highest peak in the McDonnells, Mount Giles) is Australia’s 3rd highest mountain above sea level, at 1,389m.The Ochre Pits is an Aboriginal sacred site. The rock walls are covered with layers of red, yellow and white ochre.It provides material traditionally uised for ceremonies and trading.Ormiston Gorge is 136 km from Alice Springs and contains a quite large semi-permanent waterhole. A great place for a swim on a hot day.Felix and I at Ormiston Gorge
We drove a little further on from Ormiston Gorge to the Finke River crossing, then turned back to head to Alice Springs, stopping on the way at Serpentine Gorge and Ellery Creek Big Hole. I have to say, despite the spectacular beauty of the places we had already visited, I was blown away by Serpentine Gorge. We climbed a steep path to the top of the eastern wall of the gorge, and the views in all directions were stunning. Felix and I saw another rare Black-Footed Wallaby quite close to us on a rock near the rim of the cliff.
Serpentine GorgeView south from Serpentine GorgeMe on the rim of the gorge, and the view towards the west.Ellery Creek Big HoleEllery Creek Big Hole
To be continued…..as we head south west towards Kings Canyon.
Does lubrication of the TKR by synovial fluid need to be taken into account in estimating wear?
In Part 2 of this series (see here), I examined whether powerlifting would significantly reduce the lifetime of a total knee replacement (TKR). I used Archard’s Law to do some analysis of the additional wear associated with the increased load on the joint of typical powerlifting training. Archard’s Law assumes a constant wear coefficient for the polyethylene bearing in the TKR so that the polyethylene wear rate is proportional to the load on the joint.
A reader asked me whether researchers have tried to model polyethylene wear rates, taking into account any lubrication effect that the body’s synovial fluids might provide to the prosthetic joints? If synovial fluids act as a true lubricant to prevent wear contact between the surfaces, then Archard’s Law may not apply.
How lubrication affects wear
Synovial fluids are complex fluids responsible for the lubrication present in our joints. These fluids consist of various constituents, including hyaluronic acid, surface-active proteins (i.e., lubricin), surface-active phospholipids, as well as various other proteins, each of them playing an essential role in lubrication.
Kung et al. (2015) reviewed 11 papers that examined synovial fluids providing lubrication to knee prostheses after joint replacement. They found that the cells in the fluid were similar to those in normal joints, protein and phospholipid concentrations were similar, but that the synovial fluids around joint replacement devices were typically lower in viscosity than pre-arthroplasty fluids. They concluded that the lubricant formed after joint replacement was adequate for good performance in most cases, but that studies were needed on its role in component wear or failure.
The primary purpose of lubrication is to create an acceptable lubricant film to sufficiently keep the two moving surfaces apart while allowing them to move with reduced friction. This is the ideal condition, but a lubricant can pass through several different regimes before it achieves this full film format. These regimes are associated with different frictional levels and can effectively result in non-linear changes in wear as a function of load (Mang and Dresel 2007, Maru and Tanaka 2007).
Lubrication works by separating surfaces with a film of fluid, but in boundary and mixed lubrication regimes, there are still contact points called asperities. When the load increases, these asperities come into greater contact, leading to increased wear and adhesion.
The effective distance between the surfaces is the main parameter that influences lubricated wear behaviour. This distance usually is measured in terms of the ratio between the effective distance between the surfaces and the root mean square of roughness height (λ). In machines, the main factors that reduce λ are increased load or surface roughness and reduced relative speed or lubricant viscosity.
The following Figure illustrates the relation between the friction coefficient (μ), the wear coefficient (k), and λ. Values of λ greater than 3 results in little or no solid contact, which causes negligible wear and increased wear and adhesion. If changes in load result in transition across any of these regimes, this can effectively result in non-linear changes in wear rate as a function of load.
Figure 1. The Stribeck curves shows how friction and lubrication regimes change with load. Four different regimes can be identified for lubricated contacts: hydrodynamic; elasto-hydrodynamic (EHL); mixed; and boundary. In the boundary lubrication phase, which occurs at higher loads or in thinner films, the wear coefficient tends to increase. . Hydrodynamic lubrication results in increased separation, reducing contact and thus wear (Mang and Dresel 2007).
Increasing the load on sliding surfaces should reduce the thickness of the lubricant layer and increase the wear coefficient. However, I came across one paper which carried out experiments with lubricated metal-metal contact and found that the wear coefficient effectively decreased as load increased so that wear was less than predicted by Archard Law’s proportionality to load (Ideris et al 2023).
Modelling the impact of synovial fluid lubrication on prosthesis wear rates
As for the modelling of the effect of synovial fluid lubrication in reducing wear in a TKR, researchers are only beginning to grapple with the challenges of determining the impact of synovial fluids on hip and knee prosthesis wear (Marian et al 2021, Gao et al 2022)
However, the analysis I performed in my previous article used observed failure rates of TKRs and such failure rates already include whatever impact synovial fluids have in lubricating the prostheses. Where the simplistic analysis I made using Archard’s Law may go wrong is in its assumption of a constant wear coefficient – meaning that that wear is proportional to load according to Archard’s Law. If changes in the lubricants film thickness or flow characteristics are associated with increased load, it possible that the additional wear associated with the additional load during powerlifting training may be greater than that estimated assuming its proportional to total load.
Potential impact of powerlifting on prosthesis failure rates
My previous calculations estimated that my typical training regime might result in a 1-2% increase in wear debris overall, and this would correspond to a 1-2% increase in the revision rate at 20 years from 8.7% to 8.9%. Suppose that deviations from Archard’s Law associated with synovial lubrication resulted in the additional powerlifting load adding double or triple that percentage to wear debris. The revision rate at 20 years might rise from 8.7% to 9.0 or 9.2%.
We also need to factor in the continuing improvements in prosthesis failure rates. The failure rates I used in my previous calculations came from an analysis of data from national joint replacement registries and the 20-year failure rates would have mainly related to TKRs carried out in the late 1990s. My prostheses implanted in 2022 and 2025 almost certainly have lower average failure rates.
The most recent data I could find from the Australian national registry had 20-year revision rates of 7.7%, 7.6% and 8% in 2023, 2024 and 2025 (Smith et al 2023, Lewis et al 2024, Lewis et al 2025). I think its reasonable to assume that improvements in failure rates for prostheses of the 2020s compared to those of the early 2000s have more than outweighed any increase in failure rate associated with powerlifting training.
Pending improved data on the relationship of prosthesis wear to load, I stick by my conclusions in the previous article. “Powerlifting training after joint replacements by an experienced lifter with attention to technique and careful progression appears unlikely to significantly decrease hip or knee replacement lifetimes. Indeed, the improvements in muscular strength around these joints from training may result in less forces acting in the joint across all activities and more than offset the effects of higher loads on wear.
My second total knee replacement and rehabilitation
I had my second TKR of the right knee just over four months ago on 2 September 2025. I saw a physiotherapist twice a week for three months and improved my maximum knee flexion to 140 degrees. My left knee, three and a half years after TKR, has a maximum flexion of 145 degrees. However, I had some pain in flexing my right knee through 90 degrees which took close to four months to fully go away.
Just a week ago, I did a walk in the French mountains to the east of Geneva with my son. It had snowed the night before and we walked 14 kilometres involving an 800-metre ascent and descent on sometimes steep and icy trails. I had no pain whatsoever in either knee and felt like I was definitely back to good functioning, even if needing a little more cardiovascular endurance. I include a few photos below.
At the three-month mark, I started powerlifting training again. Initially with an empty bar, and then a slow and steady progression to build strength not only in the muscles but also in the tendons and ligaments. Currently I am deadlifting 100 kg for sets of five and squatting below parallel with 70 kg for sets of five. Today I tested what I could lift for 1 rep, achieving 90 kg for the squat and 130 kg for the deadlift. If my training continues to go well, I hope to compete in late March, aiming for squat around 100 kg or more and a deadlift around 160 kg or more.
Walking on Les Voirons in winterMe admiring the fresh snow on the treesThe view from the summit ridge towards the Alps. Mont Blanc (4820m) is the high peak in the middle. Ten years ago I climbed it via the ridge visible in the middle, with the sun on one side.My dog Loki, the Dog of Mischief, admiring the view.
REFERENCES
Gao L, Lu X, Zhang X, Meng Q, Jin Z (2022). Lubrication Modelling of Artificial Joint Replacements: Current Status and Future Challenges. Lubricants. 2022; 10(10):238. https://doi.org/10.3390/lubricants10100238
Ideris M, Kamaruddin S, Sulaiman M, Sukindar NA, Azhar A, Yasir A. (2023). Effects of Coating and Lubrication on Friction and Wear for Metal-to Metal Application. Journal of Advanced Research in Applied Mechanics. 110. 52-62. 10.37934/aram.110.1.5262.
Kung MS, Markantonis J, Nelson SD, Campbell P (2015). The Synovial Lining and Synovial Fluid Properties after Joint Arthroplasty. Lubricants. 2015; 3(2):394-412. https://doi.org/10.3390/lubricants3020394
Marian M, Shah R, Gashi B, Zhang S, Bhavnani K, Wartzack S, Rosenkranz A (2021), Exploring the lubrication mechanisms of synovial fluids for joint longevity – A perspective, Colloids and Surfaces B: Biointerfaces, Volume 206, 2021, 111926, https://doi.org/10.1016/j.colsurfb.2021.111926.
Maru MM, Tanaka, DK (2007). Consideration of stribeck diagram parameters in the investigation on wear and friction behavior in lubricated sliding. Journal of the Brazilian Society of Mechanical Sciences and Engineering, 29(1). https://doi.org/10.1590/s1678-58782007000100009
Lewis PL, Gill DR, McAuliffe MJ, McDougall C, Stoney JD, Vertullo CJ, Wall CJ, Corfield S, Du P, Holder C, Harries D, Edwards S, Xu A, Lorimer MF, Cashman K, Smith PN (2024). Hip, Knee and Shoulder Arthroplasty: 2024 Annual Report, Australian Orthopaedic Association National Joint Replacement Registry, AOA: Adelaide, South Australia. 2024. https://doi.org/10.25310/GLOL7776
Lewis PL, Gill DR, McAuliffe MJ, Stoney JD, Vertullo CJ, Wall CJ, Corfield S, Esaian R, Moylan S, Du P, Holder C, Edwards S, Xu Q, Oakey H, Lorimer MF, Smith PN (2025). Hip, Knee and Shoulder Arthroplasty: 2025 Annual Report, Australian Orthopaedic Association National Joint Replacement Registry, AOA: Adelaide, South Australia. 2025. https://doi.org/10.25310/MXFR3061
Smith PN, Gill DR, McAuliffe MJ, McDougall C, Stoney JD, Vertullo CJ, Wall CJ, Corfield S, Page R, Cuthbert AR, Du P, Harries D, Holder C, Lorimer MF, Cashman K, Lewis PL (2023). Hip, Knee and Shoulder Arthroplasty: 2023 Annual Report, Australian Orthopaedic Association National Joint Replacement Registry, AOA: Adelaide, South Australia. 2023. https://doi.org/10.25310/YWQZ9375