A recently published paper in Nature (Oct 18) has analysed the mitochondrial DNA of 1,200 indigenous Africans living in the southern part of Africa and identified the ancestral homeland of all humans alive today, the place where mitochondrial Eve lived nearly 200,000 years ago. More on that below, but first some background.
In February 2014, I did a series of posts on my deep maternal ancestors, identified through a test of mutations on my mitochondrial DNA (mtDNA) which is inherited only from the mother. These mutations allowed me to track back through time to mitochondrial Eve, the single woman from whom all humans alive today descended through their female line (mother to mother to mother….). Specific mutations on the mtDNA define maternal haplogroups, and the founder of a given haplogroups is the specific individual woman in which the defining mutation occurred. All members of a given haplogroup trace their maternal ancestry back to this founder.
DNA tests have become much less expensive, and can include much more detailed testing. In the last three months, I’ve redone a test on my mtDNA and also on my Y DNA, which is inherited only down the male line (father to father to father….). I am still digesting the results of these tests, and will post on them in the near future. One of the first things I discovered was that the dates associated with haplogroup founders have been revised over time, and as more and more test results are available, and that the terminology used for identifying haplogroups has also evolved. I also came across very recent research which has pinned down the location where mitochondrial Eve lived, as well as revised estimates of the time period in which she lived.
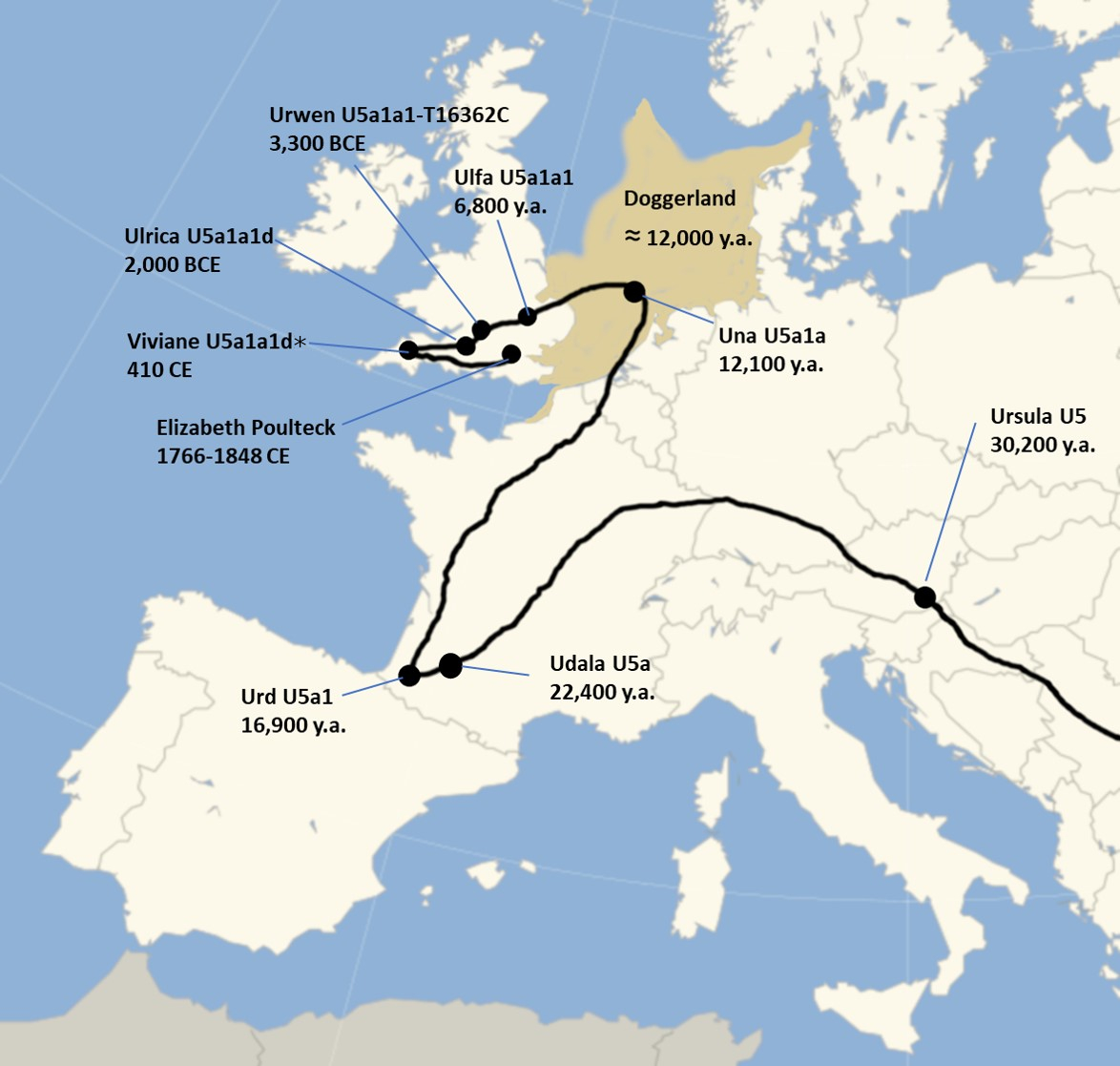
Haplogroup U5 – the oldest of seven native European haplogroups
My mtDNA haplogroup is U5, the oldest of the seven native European haplogroups. Haplogroup U most likely arose in the Near East, and spread into Europe in a very early expansion, giving rise to seven native European haplogroups, including U5. The presence of haplogroup U5 in Europe pre-dates the last ice age and the expansion of agriculture in Europe. Today, about 10% of modern Europeans are the direct maternal descendants of the founder U5 woman, who has been given the nickname Ursula*. They are particularly well represented in western Britain and Scandinavia.
Ancestral migration path of maternal ancestors for haplogroup U5
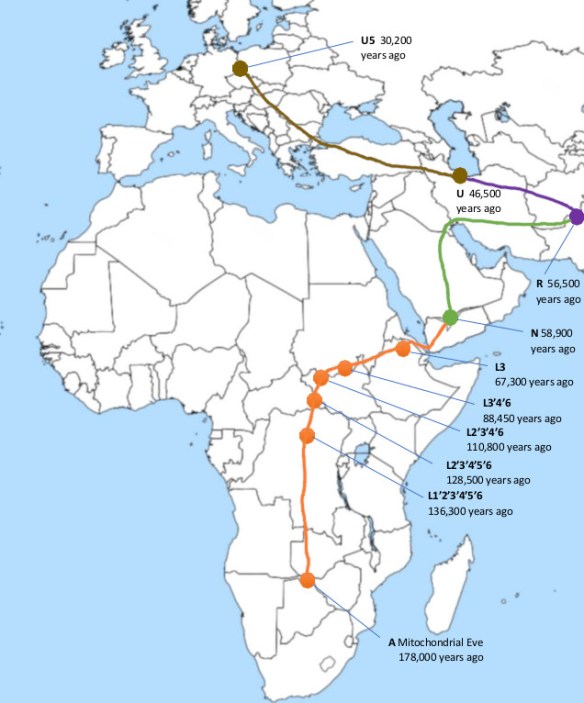
Haplogroup U in turn is descended via haplogroups R and N from haplogroup L3, which is associated with a migration of humans out of Africa around 70,000 to 50,000 years ago. The dominant theory of human origins, the “recent African origin” theory, proposes that all modern non-African populations are substantially descended from populations of H. sapiens that left Africa after during that time period. H. sapiens most likely developed in Africa between 300,000 and 200,000 years ago, and there were at least several “out-of-Africa” migrations of modern humans, possibly beginning as early as 270,000 years ago. These early dispersals may have died out or retreated, although some paleoanthropologists argue that they possibly interbred with various other local hominid species and with later humans from “recent-out-Africa” and it just so happens that all the maternal lineages trace back to “recent-out-Africa”. Of all the lineages present in Africa, only the female descendants of Lara*, founder of the L3 haplogroup, are found outside Africa. If there had been several migrations, one would expect descendants of more than one lineage to be found. Of course, all this could be upturned if descendants of other African lineages are found outside Africa, and can be traced back to earlier migrations.
Mitochondrial Eve (haplogroup L)
Mitochondrial Eve (mt-Eve) is a member of Haplogroup L and lived just before the  divergence of macro-haplogroup L into L0 and L1–6 (see diagram below). Today the haplogroup L0 and its offshoots are found mainly in southern and eastern Africa, with particularly high frequencies among the San people (bushmen) of Botswana, Namibia and other countries of southern Africa.
divergence of macro-haplogroup L into L0 and L1–6 (see diagram below). Today the haplogroup L0 and its offshoots are found mainly in southern and eastern Africa, with particularly high frequencies among the San people (bushmen) of Botswana, Namibia and other countries of southern Africa.
Haplogroup L1 is found in West and Central sub-Saharan Africa. The descendants of haplogroup L1 are also African haplogroups L2 and L3, the latter of which gave rise to all non-African haplogroups.
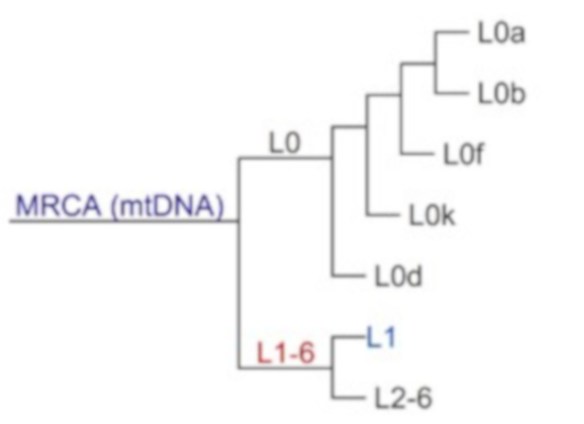
Phylogenetic tree for mtDNA Haplogroup L, commencing with mitochondrial Eve, the most recent common maternal ancestor (MRCA) of all humans.
A recent paper by Chan et al. in Nature (October 2019) [1] analysed the genomes of more than 1,200 indigenous Africans living in southern Africa and claim to have identified precisely where and when the L haplogroup split into L0 and L1 and when these groups migrated from their homeland.
Chan et al. identified this homeland as Makgadikgadi, a vast wetland some 120,000 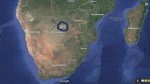 square kilometers in area, or roughly twice the area of Lake Victoria, Africa’s largest lake today. Mitochondrial Eve and her descendants lived in this region for about 30,000 years (from 200,000 to 170,000 years ago) before the L0 lineage split into its first subgroup. Today, Makgadikgadi is one of the largest salt flats in the world. Climate models suggest that, 200,000 years ago, it was a fertile oasis. The map shows the overall location of Makgadikgadi in southern Africa, and the following map shows a more detailed view.
square kilometers in area, or roughly twice the area of Lake Victoria, Africa’s largest lake today. Mitochondrial Eve and her descendants lived in this region for about 30,000 years (from 200,000 to 170,000 years ago) before the L0 lineage split into its first subgroup. Today, Makgadikgadi is one of the largest salt flats in the world. Climate models suggest that, 200,000 years ago, it was a fertile oasis. The map shows the overall location of Makgadikgadi in southern Africa, and the following map shows a more detailed view.
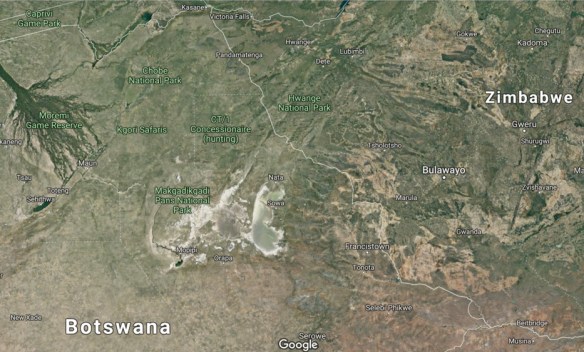
Satellite view of the Makgadikgadi salt pans. This area is located about 250 km south of Victoria Falls close to the borders of Zambia, Zimbabwe and Botswana.
Chan et al [1] date the deepest rooting L0 branch to 200,000 years ago (with 95% confidence interval 165,000 – 240,000 years ago). I have reviewed the most recent comprehensive dating of maternal haplogroups and found that the dates in Fu et al (2012) [2] and Behar et al [2013] were in reasonably good agreement. I have used dates from Behar et al, which give a date of 176,700 years ago (confidence interval ± 11,300 years) for mitochondrial Eve, and 136,300 (± 11,700) years ago for L1. This is substantially earlier than the date of the recent out-of-Africa dispersal of L3 around 65,000 years ago.
Migrations from the Makgadikgadi homeland
The Makgadikgadi wetlands were large, wet, and lush with vegetation. They would have provided an ideal home for wildlife and for early humans, including mt-Eve. So why did some migrate? Around 130,000 years ago, there was a major climatic shift associated with the end of the penultimate glacial period. This led to higher rainfall and created “green corridors” leading to the northeast and to the southwest. In particular, it appears that the ancestral founder of the L1 haplogroup lived around 136,000 years ago among a group that had migrated north into Zambia, and by around 70,000 years ago her descendents had made their way north to the horn of Africa, where Lara (L3 haplogroup founder live).
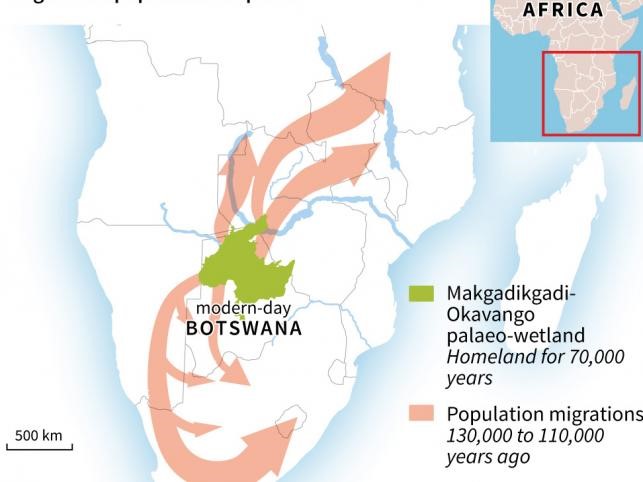
The “green corridors” proposed by Chan et al [1] helped lead humans out of the ancestral homeland
Chan and his group have extrapolated the likely location of mt-Eve’s homeland from the present-day distribution of the L haplgroup in Southern Africa, and it is always possible that future data may lead to revisions of this conclusion. However, multiple sets of evidence lead to the conclusion that mt-Eve was among the ancestors of the San people of southern Africa, although of course we likely will never know for sure exactly where she lived. And this was not the only ancestral human homeland. Y-DNA evidence suggests that Y-Adam lived in West Africa in a time period even further in the past (this will be subject of a future post) and of course, there may be other ancestral homelands associated with the many other ancestral lines than the purely maternal and paternal.

The San people of southern Africa have one of the most oldest maternal DNA lineages on Earth. They share the Haplogroup L with mitochondrial Eve who lived in northern Botswana nearly 200,000 years ago.
* Bryan Sykes in his 2001 book The seven daughters of Eve gave names to each of the women who founded the seven native European haplogroups, and also names to some of their ancestral haplogroups. He chose names that began with the letter by which the haplogroup was identified. Oppenheimer (The Origins of the British: A Genetic Detective Story, 2006) followed this example and also gave names to both mtDNA and Y-DNA haplogroups. To emphasise that the maternal clan founders were real individuals, who were my ancesters, I have used these names and given my own names to the more recent subgroup founders.
References
[1] Chan EKF, Hardie RA, Petersen DC, Beeson K, Bornman RMS, et al. (2015) Revised Timeline and Distribution of the Earliest Diverged Human Maternal Lineages in Southern Africa. PLOS ONE 10(3): e0121223.
https://doi.org/10.1371/journal.pone.0121223
[2] Fu Q, Mittnik A, Johnson PLF, et al. A revised timescale for human evolution based on ancient mitochondrial genomes. Curr Biol. 2013;23(7):553–559. doi:10.1016/j.cub.2013.02.044
https://www.cell.com/current-biology/fulltext/S0960-9822(13)00215-7?code=cell-site
[3] Behar D, van Oven M, Rosset S, et al. A “Copernican” Reassessment of the Human Mitochondrial DNA Tree from Its Root. Am J Hum Genet. 2012;90(5):936. doi:10.1016/j.ajhg.2012.04.007
Open ArchiveDOI:https://doi.org/10.1016/j.ajhg.2012.03.002

 There is an immense amount of recent research on European Bronze Age genetics and migrations and a very considerable unfinished debate on how to interpret the evidence. New evidence is being published almost weekly, as increasing numbers of ancient European remains are analysed and Y haplogroup identified. So this post will only aim to describe the big picture as best I can summarize it, and details and perhaps even some of the big picture will likely change in the future.
There is an immense amount of recent research on European Bronze Age genetics and migrations and a very considerable unfinished debate on how to interpret the evidence. New evidence is being published almost weekly, as increasing numbers of ancient European remains are analysed and Y haplogroup identified. So this post will only aim to describe the big picture as best I can summarize it, and details and perhaps even some of the big picture will likely change in the future.